







1. Introduction
Prum et al.'s study employed targeted next-generation DNA sequencing to deeply analyze the evolutionary relationships of birds using molecular data [1]. They conducted a comprehensive phylogenetic analysis of 198 bird species worldwide, estimating that the total number of extant bird species exceeds 10,000. In contrast, Jetz et al.'s research combined multi-genome analysis methods to comprehensively assess the spatial and temporal distribution of living birds [2]. Their paper estimates the global bird species count to be approximately 9,993. The diversification rate changes across the avian phylogenetic tree are shown in figure 1, where the branches represent different bird lineages, helping trace the evolutionary history of modern birds. This figure provides insight into the development of species diversity among modern birds (see figure 1). For instance, the structure of bird beaks is closely related to their foraging behavior across different avian groups. For instance, raptors within the orders Accipitriformes and Falconiformes possess powerful hooked beaks and sharp talons, which are well-suited for predation on other birds, small mammals, reptiles, and fish [3]. In contrast, the beaks of Anseriformes have evolved into flat shapes, equipped with filtering structures, making them adept at foraging and filtering food in aquatic environments. Passeriformes exhibit even greater diversity in beak morphology; some species have short, conical beaks, ideal for crushing seed coats and consuming seeds, while others have long, pointed beaks specialized for capturing insects [4]. In terms of nesting behavior, birds demonstrate remarkable creativity and ecological adaptability. Woodpeckers from the order Piciformes use their strong beaks to excavate cavities in tree trunks, showcasing a high degree of adaptation to woody environments. Conversely, certain swallows within Passeriformes, such as the barn swallow (Hirundo rustica), build cup-shaped nests on vertical surfaces using a mixture of saliva and mud [5]. On the other hand, birds in the family Cuculidae, such as the Oriental cuckoo (Cuculus poliocephalus) and the common cuckoo (Cuculus canorus), exhibit brood parasitism, laying their eggs in the nests of other bird species. Their chicks have shorter incubation periods and display aggressive behaviors, such as killing the host's chicks, to ensure their own survival. Avian courtship and reproductive behaviors are equally diverse and complex [6]. Male peafowl (Pavo cristatus) perform visual displays by fanning out their elaborate tail feathers to attract females, a behavior characteristic of sexual selection. In contrast, albatrosses (family Diomedeidae) engage in intricate and prolonged courtship dances to establish and maintain monogamous pair bonds, reflecting their sociality and the adaptive significance of long-term partnerships.
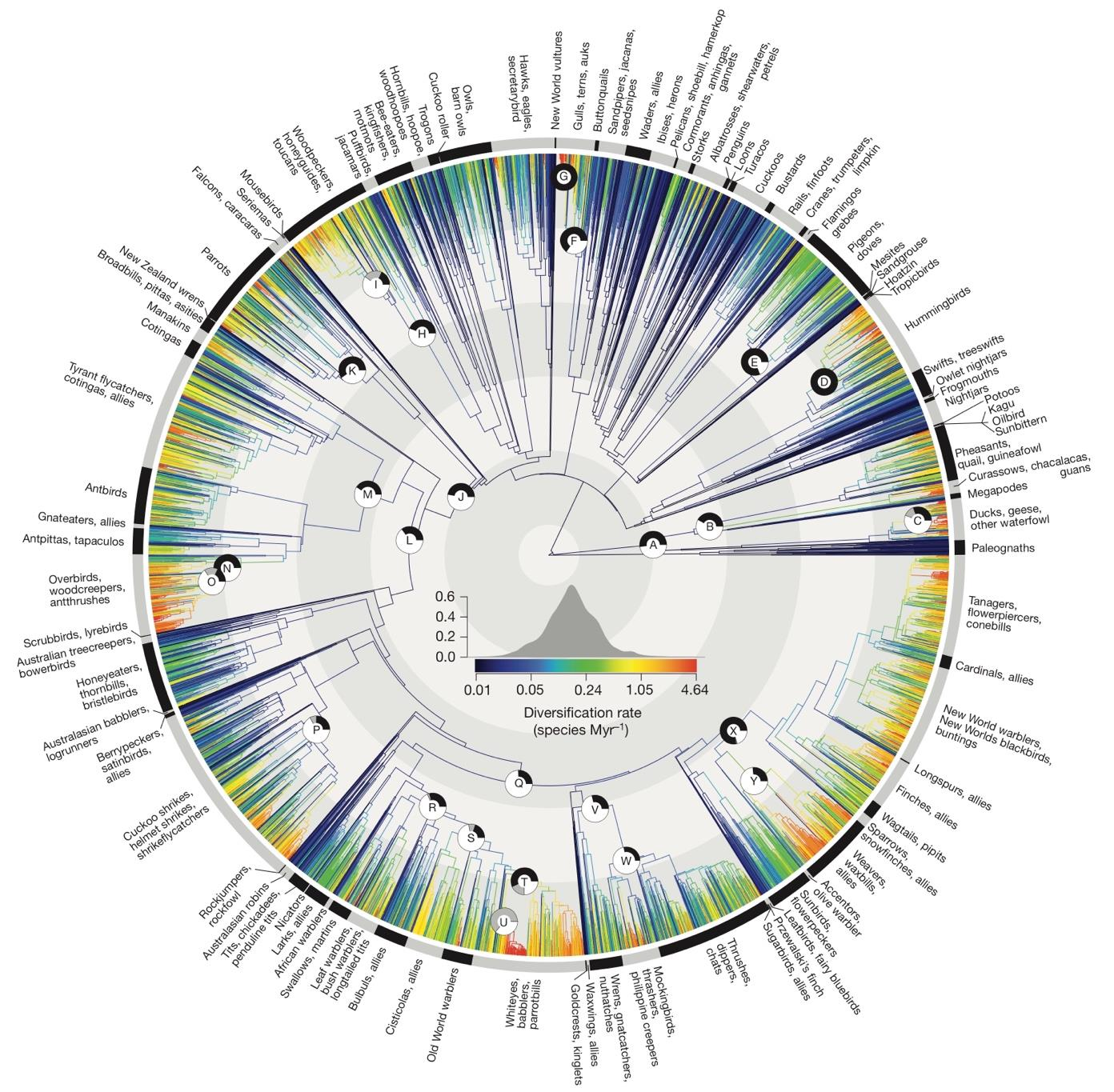
Figure 1. The avian phylogenetic tree demonstrates the extensive diversification of bird species, illustrating that global bird diversity exceeds 10,000 species. Significant diversification shifts are indicated by labeled nodes [2].
The diversity in birds is not only a result of biological evolution but also a direct demonstration of natural selection and niche differentiation. Birds have adapted to nearly all types of habitats on Earth, ranging from tropical rainforests to deserts, and from high mountains to oceans [7]. However, this diversity is not evenly distributed across the globe, with the highest concentrations in tropical regions of South America, Southeast Asia, and Africa. Their numbers are influenced by a variety of factors including habitat diversity and climatic conditions [8]. The species diversity and thriving populations of birds represent healthy ecosystems because birds occupy different ecological niches, including herbivores, predators, and scavengers, which reflect the abundance of primary productivity [9]. An ecosystem with an abundance of birds and a healthy population size is usually indicative of good energy flow and material cycling within the ecosystem. Changes in bird populations are often an early warning of environmental stresses on ecosystem health, such as climate change, habitat loss or habitat pollution [10]. For example, habitat destruction (including both pollution and fragmentation) can reduce the numbers of arthropods, amphibians, reptiles, and small mammals, subsequently affecting the availability of prey for birds, thus reducing their population numbers, while climate change can alter habitats and food availability, leading to shifts in migration patterns, breeding success, and ultimately causing declines in bird populations.
Birds play a crucial role in maintaining ecological balance [9]. They assist in vegetation recovery through seed dispersal, control pest populations to protect crops, and positively affect agricultural and forestry developments. Birds also connect multiple levels of the food chain, such as preying on insects, small mammals, or other birds. Additionally, the migratory behavior of birds facilitates cross-regional exchanges of materials and energy, underscoring their ecological value. Thriving bird populations more effectively fulfill these roles, thereby supporting the overall health and stability of ecosystems. The effective performance of these services relies on maintaining avian diversity, as each bird species has its unique role and function. As high-level consumers, the diversity of birds reflects the complexity and productivity of the lower biological communities. A diverse and thriving bird population generally indicates a high degree of biodiversity and a well-balanced ecosystem in its habitat.
Birds also hold significant symbolic value in many cultures, and protecting them means safeguarding culture and inspiration [11]. These birds serve as valuable tools for natural education and scientific outreach. In various cultures, birds often symbolize concepts such as freedom, peace, or rebirth. For example, the Rock Pigeon (Columba livia) is globally recognized as a symbol of peace, while the Red-crowned Crane (Grus japonensis) in Chinese culture symbolizes longevity and grace. These symbolic meanings make birds a vital part of cultural heritage. Protecting birds also protects these deeply rooted cultural symbols and traditional stories, crucial for maintaining cultural diversity. Furthermore, birds, with their unique behaviors, beautiful plumage, and melodious songs, often inspire artists and writers. Many famous poems, paintings, and musical compositions are inspired by birds. Protecting birds preserves these sources of inspiration, providing continuous material for future artistic endeavors. Birds, as easily observable wildlife, are excellent resources for natural education. Observing birds allows people to learn about biodiversity, ecosystem functions, and their interactions. Birdwatching activities in schools and communities can raise public awareness about biological conservation [12]. In addition, birdwatching, as a form of ecotourism, attracts millions of enthusiasts globally. This not only enhances public awareness of nature conservation but also directly stimulates local economic development. The income generated from birdwatching provides economic incentives for many regions to protect natural environments.
Birds are an indispensable group within ecosystems, because of their sensitivity, they are often indicators of environmental change - the presence or absence of birds can reflect the health of an ecosystem [9]. The development of human society inevitably leads to intensified conflicts between humans and wildlife, a well-recognized fact, with one of the greatest threats to birds being habitat loss, degradation, and fragmentation caused by human activities [13]. These changes reduce the availability of essential resources needed for avian survival. Urban expansion not only encroaches on bird habitats but also poses direct threats to individual birds, such as collisions with wind turbines or buildings, electrocution by power lines, or vehicle strikes [14]. In North America alone, up to one billion birds die annually from collisions with buildings [15]. Therefore, scientifically designing buildings to reduce urban light pollution and glass reflection and creating bird-friendly cities are crucial. Bird-friendly cities serve as excellent examples of habitats shared by humans and wildlife. Coexistence strategies can protect wildlife habitats maximally without impairing normal human societal functions and living conditions.
Birds exhibit remarkable diversity in behavior, morphology, and ecological roles. Among them, corvids stand out not only for their intelligence but also for their complex social behaviors and extraordinary adaptability to various environments. The motivation for this study lies in the unique position that corvids occupy within the avian world. Their ability to thrive in both natural and urban environments, combined with their advanced cognitive functions, makes them ideal subjects for studying the impacts of environmental changes and the evolutionary processes driving avian adaptability. Understanding corvids' interactions within their social structures and their responses to human-induced environmental changes can provide critical insights into broader ecological and evolutionary dynamics.
This paper addresses the following questions, which are still fully addressed: How does the complexity of corvid social structures influence their ability to learn and transmit knowledge? How do social interactions impact their cognitive development? Additionally, how does the growing phenomenon of urbanization affect corvids' food choices and habitat use? The influence of human activities, such as feeding, on corvid behavioral adaptations and social structures is another area of interest this paper will explore. Beyond these social and behavioral aspects, the ecological roles of corvids are crucial for understanding their impact on ecosystems. This paper will examine how corvids' foraging behaviors, such as scatter hoarding, influence plant population distribution and ecosystem structure, as well as the specific differences in scavenging behaviors across various environments and their ecological significance. Finally, it will explore how corvids adapt to diverse ecological environments through genetic variation and whether niche differentiation drives significant genetic divergence within corvid populations.
2. Comprehensive Analysis of Corvid Intelligence, Social Behavior, and Ecological Adaptability Across Diverse Environments
Corvids, including species such as crows, ravens, jays and magpies, are widely distributed across the globe, with their presence felt almost everywhere except for the extreme regions of South America and the polar areas. These birds are renowned for their remarkable intelligence and problem-solving abilities, making them considered among the most intelligent of all bird groups. Corvids can be commonly found in our surroundings (See figure 2).



Figure 2. Corvids observed during personal fieldwork. The species shown from left to right are: Steller's Jay (Cyanocitta stelleri) coastal morphotype C. s. carbonacea (left image, photographed on October 12, 2024, in Vancouver, BC, Canada); Eurasian Magpie (Pica pica), middle image, photographed in August 2024 in Beijing, China; Azure-winged Magpie (Cyanopica cyanus), right image, showing a juvenile born this year, photographed in July 2024 in Shaanxi Province, China. All images were captured during my personal observations.
Corvid social behavior is equally complex. They are capable of displaying cooperative behaviors, such as jointly defending territories or collectively caring for offspring, and exhibit clear hierarchical structures and social organization within their groups. These birds can even express complex social emotions, such as a sense of fairness and deceit, which require sophisticated social cognitive abilities.
In the realm of cognition, corvids exhibit advanced cognitive functions similar to those of humans and other higher animals. Eurasian Magpie (Pica pica) can recognize themselves in a mirror during the self-recognition test, indicating that they have the ability of visual self-recognition [16]. For instance, crows can craft tools from natural materials like branches to access food that is otherwise unreachable, demonstrating their innate ability to utilize tools. Moreover, these birds are capable of understanding cause-and-effect relationships, engaging in future planning, and exhibiting strategic thinking, such as navigating complex multi-step processes to unlock food from containers. These behaviors not only showcase their intelligence but also reveal the highly developed abstract thinking and problem-solving capacities within their brains.
In terms of environmental adaptation, corvids display remarkable flexibility. Their diet is extensive, ranging from invertebrates to small mammals, and includes fruits, seeds, and even carrion. This omnivorous nature allows them to adjust their feeding habits in response to changes in available resources, adapting to various survival conditions. Amidst urbanization and climate change, corvids adapt by altering their range of activity and habitat selection, demonstrating exceptional ecological adaptability.
Furthermore, the contribution of corvids to ecosystem services is significant. Their behavior of scatter-hoarding aids in forest regeneration, as caching seeds in the soil, is crucial for the reproduction and dispersal of certain tree species. Their presence helps maintain the health and stability of ecosystems, managing pest populations and cleaning up decaying matter, thus preserving ecological balance.
2.1. The Influence of Corvids' Complex Social Structures on Their Capacity for Learning and Knowledge Transmission
Corvids are renowned for their highly developed social structures and cognitive capabilities. These birds exhibit remarkable learning and knowledge transmission abilities through complex social interactions. The complexity of corvid social structures significantly influences their cognitive development and learning abilities, primarily manifested in the dynamics of group living, including social learning, information sharing, and problem-solving strategies.
Corvids possess exceptional social learning capabilities, as validated by numerous studies. For instance, research on the California Scrub-Jay (Aphelocoma californica) indicates that individuals from larger groups perform better in cognitive tasks [17]. These findings suggest that the frequency and complexity of social interactions within the group directly impact an individual's cognitive abilities. Larger groups provide more social information and relationships, thereby facilitating the development of memory and problem-solving skills and enhancing individual adaptability in complex environments.
Social experiences, particularly those during juvenile stages, are crucial for the social interactions and cognitive development of corvids [18]. Social experiences largely determine these birds' patterns of social behavior. Early social interactions shape their future social behaviors and cognitive skills. For example, the New Caledonian Crow (Corvus moneduloides) utilizes tools like branches to obtain food, a capability learned through observing and mimicking the behavior of other group members [16]. This type of social learning is not confined to kinship members but also extends to interactions with non-kin group members, further enhancing learning efficiency.
Research on corvids also highlights the importance of non-breeding social interactions for understanding avian social complexity. Social interactions and challenges faced by Common Ravens (Corvus corax) during the non-breeding season, such as food competition and social status contests, significantly influence their cognitive performance [19]. Ravens exhibit high levels of strategical and adaptive responses to these social challenges, which drive their cognitive evolution. Behaviors such as hiding food to prevent theft by conspecifics not only demonstrate their memory capabilities but also reflect their ability to reason about the behaviors of their peers.
2.2. The Impact of Urbanization and Human Activities on the Behavioral and Social Structure of Corvids
In this section, the paper emphasizes the impact of ecological niche changes on the behavior of corvids. Initially, it addresses how urbanization influences the food choices and habitat use of corvids. The process of urbanization provides corvids with abundant and diverse food resources and habitat options. Research indicates that many corvids utilize man-made structures such as buildings and poles for nesting [20]. For instance, the nesting behavior of the Hooded Crow (Corvus cornix) and Azure-winged Magpie (Cyanopica cyanus) in urban areas effectively utilizes buildings and utility poles. These artificial structures not only provide safe habitats but also reduce predation threats. Moreover, urban waste, food remnants, and human feeding practices, such as the placement of bird feeders, offer stable and reliable food sources for corvids [21]. The richness of these resources enables the urban populations of corvids to maintain and increase, especially during the food-scarce winter months [22]. In winter, corvids exhibit selectivity for specific habitat features. Studies have found that green spaces and blue infrastructure, such as rivers and lakes, are crucial habitats for corvids. For example, the Eurasian Jay (Garrulus glandarius) prefers areas with tree cover, while the Eurasian Magpie and the Carrion Crow (Corvus corone) tend to choose more urbanized areas [23]. This selectivity is not only related to the distribution of food resources but also to the availability of shelters and the degree of human activity interference.
The second focus is on how human activities in urban environments affect the behavioral adaptations and social structures of corvids. Human feeding behavior is particularly common in urban areas, especially during winter. This practice significantly reduces the risk of starvation for birds, helping them survive the cold seasons. Studies show that urban waste and food remnants near restaurants also become significant food sources for corvids, enhancing their survival rates and facilitating population expansion [21, 23]. Corvids demonstrate a high degree of behavioral adaptability. For example, many corvids in urban areas show high tolerance for human activities, adapting to noise and light pollution [22]. They also adapt to new environments through social learning, learning how to utilize new resources and cope with potential threats. This behavioral adaptation not only helps them survive in urban settings but also promotes their reproductive success. In urban environments, the social structure of corvids also changes. They often form large-scale flocks, such as mixed foraging groups, to enhance foraging efficiency [23]. This flocking behavior not only aids in food acquisition but also reduces predation risks to some extent. Furthermore, flocking behavior promotes information transfer and social learning, allowing individuals to quickly adapt to environmental changes [22].
Urbanization leads to the urban heat island effect, where city temperatures are typically higher than those in surrounding rural areas, particularly benefiting corvids during winter [21]. Higher temperatures and reduced snow cover make foraging easier, helping birds conserve energy. Studies indicate that as the climate warms, more corvids choose to winter in cities, a phenomenon particularly evident in species like the Eurasian Magpie and the Hooded Crow [22]. Climate change and urbanization jointly affect the population dynamics of corvids. For instance, studies in northern latitudes show that with warming climates and advancing urbanization, corvid populations during winter significantly increase. This change is not only related to the availability of food resources but also closely linked to the safe habitats provided by cities [21].
Corvids exhibit high adaptability in urban living, utilizing the rich resources and habitats provided by cities [22]. For example, they show adaptive behaviors such as reduced flight initiation distance and quickly adapt to new environments and resources through social learning. Additionally, corvids display high cognitive abilities, able to solve complex problems, which enables them to thrive in urban settings. In urban environments, there is significant habitat overlap among different corvid species. These birds typically have broad ecological niches, allowing them to coexist in the same environment. For example, the habitats of the Eurasian Magpie and the Hooded Crow highly overlap, but they reduce competition through different foraging behaviors and food preferences [20]. This niche differentiation not only reduces interspecies competition but also promotes population stability and diversity.
2.3. The Impact of Corvids on Ecosystem Services through Scatter-hoarding and Scavenging Behavior
Corvids significantly impact the distribution of plant populations and the structure of ecosystems through their foraging behavior, particularly scatter hoarding [16]. Corvids select, transport, and store seeds, a behavior that facilitates the dispersal of seeds away from the parent trees, enhancing plant gene flow and population expansion. For instance, plants like oaks and pines rely on corvids' scatter-hoarding behavior for seed dispersal, thereby forming new forest areas. Additionally, seeds cached by corvids at sites suitable for germination that are not retrieved can sprout and grow, further promoting forest regeneration, altering the spatial distribution of plant communities, and providing habitat and food resources for other animals, thereby promoting the overall health and stability of ecosystems.
Scavenging behavior refers to certain animals specifically seeking and consuming the carcasses or remnants of other animals. Some scavengers, such as corvids, exhibit complex social behaviors during foraging, potentially forming foraging groups to locate and share food resources. In different habitats, corvid scavenging behaviors exhibit significant variations [24]. In urban environments, due to the abundance of food resources such as waste and leftovers from human activities, corvid scavenging behavior is more active. The mild climate and reduced predation pressure in cities also promote high-density aggregation of corvids. In forests, corvids primarily decompose naturally deceased animals and plant detritus, facilitating organic decomposition and nutrient cycling. In agricultural areas, corvid scavenging behavior is related to crop residues and agricultural waste; they prey on agricultural pests, serving a biological control function, but may also cause damage to crops. Seasonal changes significantly affect corvid scavenging behavior, especially in winter when food resources are limited, making scavenging behavior more frequent. Additionally, following flooding events, an increase in corvid scavenging behavior may occur, possibly due to the increased availability of animal carcasses brought by the floods. Corvid scavenging behavior holds significant ecological importance. By decomposing animal carcasses and plant detritus, they facilitate nutrient cycling, helping to maintain the nutrient balance of ecosystems. Furthermore, scavenging behavior not only reduces the spread of pathogens and outbreaks of diseases but also by rapidly clearing carcasses, lowers the risk of pathogens in the environment. Additionally, during the scavenging process, corvids may further reduce disease transmission risks by preying on insects and larvae on the decomposing flesh.
2.4. Corvids Adapt to Diverse Ecological Environments through Genetic Variation, with Niche Differentiation Driving Significant Genetic Divergence among Populations — Take Steller's Jay (Cyanocitta stelleri) for Example
The Steller's Jays are mainly divided into three forms, namely coastal form, inland form, and Rocky Mountain form [25]. The coastal form of Steller's Jays primarily inhabits the moist coastal regions of North America's west coast. The coastal morphotype of Steller's Jay includes three subspecies: Cyanocitta stelleri stelleri, which is distributed in southwestern Alaska; Cyanocitta stelleri frontalis, found in the central and southern coastal regions of California; and Cyanocitta stelleri carbonacea, primarily distributed in parts of the coastal regions of British Columbia, including Vancouver Island and the adjacent coastal areas of the mainland. These geographic features typically have a moist maritime climate with higher rainfall, fog, and moderate temperatures. These environmental conditions provide Steller's Jays with abundant food resources and favorable nesting sites. The coastal form is larger, which helps maintain body heat in the cool coastal climate, as larger body sizes reduce heat loss per unit body surface area. Additionally, the coastal form has darker plumage, especially shorter and darker blue feathers on the forehead. Darker feathers are more effective at absorbing solar radiation, providing extra warmth in the cold coastal environment. The short, dark forehead feathers also help camouflage better in the densely forested habitat, adapting to the concealment needs of their habitat.
The interior form of Steller's Jays predominantly resides in the inland areas of North America, particularly stretching east from California to states like Nevada, Utah, and Colorado. The scientific name of this subspecies is Cyanocitta stelleri annectens. The geographic features here include more forests and grassland ecosystems, with climates ranging from semi-arid to temperate. The inland area experiences more significant seasonal variations than the coastal areas, with potentially colder winters and hotter summers, providing Steller's Jays with diverse habitat conditions. The interior form has a medium size, possibly an adaptation to the milder climate of its habitat. This medium size provides enough mobility and flexibility in the food-rich inland environment, adapting to the diverse forest and grassland areas. The plumage is lighter, with longer head feathers intermixed with blue and white, and distinct white eyebrow markings. Lighter feathers help better camouflage in the inland environment, especially against snowy or winter backgrounds, increasing their concealment capabilities. The distinct white eyebrow markings not only enhance the visibility of social interactions but may also aid in intra-species recognition and interaction.
The Rocky Mountain form of Steller's Jays mainly inhabits the Rocky Mountain region of North America, including high-altitude areas in states like Montana, Wyoming, Colorado, and New Mexico. The scientific name of this subspecies is Cyanocitta stelleri macrolopha. The geographic features include high altitude, Rocky Mountains, and open grasslands, with generally drier climate conditions and significant temperature variations, especially at night. These regions have pronounced seasonal environments, with long, cold winters and short, warm summers. The Rocky Mountain form is smaller in size, which helps them move and forage more effectively in high-altitude and rugged terrain while reducing energy demands and increasing thermal efficiency, especially in the cold mountain climate. The plumage is lighter, and suitable for the variable lighting and backgrounds of mountainous areas, helping the birds camouflage in rocky, snowy, and low-vegetation environments, thus reducing the risk of predation. The forehead feathers are very long, predominantly white, with significant white eyebrow markings. The long, conspicuous white forehead feathers play a visual signaling role in social and territorial defense behaviors, where the striking head pattern helps individuals recognize and communicate with each other from a distance in visually complex mountain environments.
The different geographical distributions of various subspecies of Steller's Jay are effectively illustrated (see figure 3). In the case of Steller's Jay, despite the Coastal, Interior, and Rocky Mountain morphotypes having multiple subspecies with different scientific names, individuals of these subspecies have undergone geographic isolation or ecological adaptation during evolution, resulting in distinct characteristics (such as body size, plumage color, etc.). However, from a biological perspective, they are still considered the same species because these subspecies can interbreed without reproductive isolation.
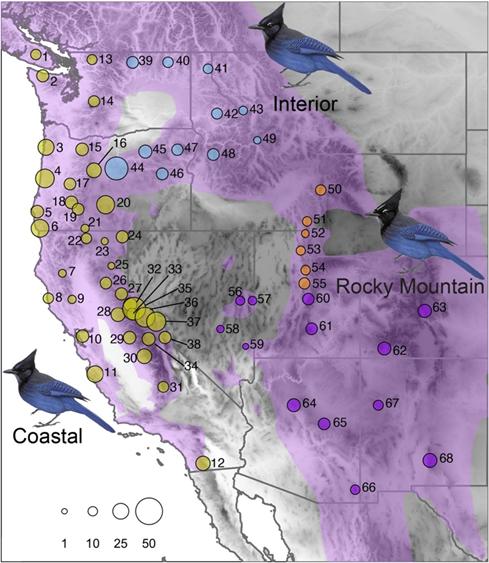
Figure 3. Geographic distribution of three distinct morphotypes of Steller's Jay (Cyanocitta stelleri): the Coastal morphotype (C. s. stelleri, C. s. frontalis, C. s. carbonacea), the Interior morphotype (C. s. annectens), and the Rocky Mountain morphotype (C. s. macrolopha). The sizes of the circles correspond to sample sizes at each location [25].
The adaptation of Steller's Jays to different ecological regions in North America showcases their evolutionary adaptability to each unique environment. Particularly, the Rocky Mountain form exhibits significant genetic divergence, likely due to long-term geographical isolation and unique selective pressures. The ecological niche model supports multiple glacial refugia, suggesting that Steller's Jays could adapt to variable historical climate conditions, with different forms being isolated during the last ice age and developing distinct adaptive traits in their respective environments. The genetic divergence between the coastal and interior forms is relatively minor, reflecting their ecological similarities, while the significant genetic differences of the Rocky Mountain form are the result of its adaptation to its unique high-altitude, dry, and open environments.
Ravkin et al. answered this question from a more general perspective. Corvids adapt to different ecological environments through genetic variation, and niche differentiation drives significant genetic divergence [26]. Different land cover types, human activities, terrain, and climate factors significantly affect the distribution and genetic variation of corvids. For instance, certain corvid species are more inclined to be active in forests and forest edges, while others are more common in open or semi-open grasslands. This diversity in land cover types, through natural selection, causes different corvid species to exhibit distinct genetic adaptations. The increase in urbanization and human activities has also had a significant impact on the distribution of corvids. In urban and suburban areas, some adaptable corvid species have increased in number because they can utilize resources available in urban environments (such as garbage), adaptability that may be achieved through genetic variations, enabling these species to better survive and reproduce in urban settings. Terrain conditions, such as mountains, plains, and river systems, similarly affect the habitat choices and foraging behaviors of corvids. For example, certain species may be more adapted to specific terrain conditions, reflected in their genes, allowing them to survive more effectively in those environments. Climate conditions, especially changes in temperature and precipitation patterns, also significantly impact the distribution of corvids. Some species may prefer cold or warm climate zones, affecting their migration and breeding activities. The influence of climate conditions on genetic variation occurs through selective pressures, making genes adapted to specific climate conditions more common in populations. Niche differentiation drives the genetic divergence of corvids through selective pressures. Corvids display different adaptive traits in different ecological niches. For example, some species excel in forest niches, while others are more common in grasslands or wetlands. This ecological niche differentiation leads to genetic divergence, as individuals in different niches experience different selective pressures, leading to the accumulation of distinct genetic variations.
3. Conclusion
This paper comprehensively explores the diversity, intelligence, social behavior, and ecological adaptability of corvids, emphasizing their unique role within avian species and broader ecosystems. With their exceptional cognitive abilities, complex social structures, and adaptability to various environments, corvids offer valuable insights into the evolutionary processes shaping avian behavior and ecology. The findings of this review highlight several key aspects of corvid life. First, the complexity of corvid social structures significantly influences their cognitive development and their ability to learn and transmit knowledge. It is evident in their advanced problem-solving abilities, tool use, and social learning behaviors, where imitation and communication among individuals enhance their survival skills and knowledge accumulation. Second, the impact of urbanization and human activities on corvids demonstrates their remarkable behavioral flexibility and ecological resilience. By altering their food choices, nesting habits, and social structures, corvids have successfully adapted to urban environments, allowing them to thrive even amidst rapid environmental changes. Third, the ecological roles of corvids, particularly in seed dispersal and scavenging, are crucial for maintaining ecosystem health and stability. Their behaviors not only contribute to forest regeneration and nutrient cycling but also help control pest populations and prevent disease transmission. Finally, this review discusses how genetic variation and niche differentiation lead to significant genetic divergence in corvid populations across different ecological environments, further emphasizing their adaptability.
The significance of this study lies in its ability to synthesize current research on corvids, providing a comprehensive understanding of how these birds navigate environmental challenges through both behavioral and genetic adaptations. By examining the interplay between social behavior, cognitive development, and ecological roles, this review contributes to a broader understanding of avian evolution and the factors driving species success across diverse habitats. This review underscores the importance of corvid conservation and, more broadly, avian conservation. The insights gained from studying corvids hold broader implications for conservation efforts, particularly in urban and rapidly changing environments. Protecting and understanding these intelligent and adaptable birds not only preserves their ecological contributions but also provides a model for how other species might adapt to the challenges posed by global environmental changes. As human activities continue to alter natural habitats, the study of corvids offers valuable lessons in resilience and adaptability, which are essential for the conservation of biodiversity in the Anthropocene.
Although corvids are among the most extensively studied bird groups, our understanding of their wide distribution and complex behaviors remains limited. If this is the case for corvids, the attention given to other less-studied avian groups is likely even more insufficient. While this study covers various environments, it may not encompass all possible habitats or geographic regions of corvids. In regions that have not been widely studied, subtle ecological and behavioral differences may exist. Additionally, this research lacks comprehensive data on some rare or less common corvid species, focusing primarily on key species, which may limit the generalizability of the findings across the entire corvid family. Behavioral studies on corvids often involve complex variables such as social dynamics, which may not be fully captured in all studies.
Future research should continue to explore the genetic and evolutionary aspects of corvids, utilizing interdisciplinary approaches that combine molecular genetics, ecology, evolutionary biology, and animal behavior to better understand how genetic factors influence the social structure and cognitive abilities of corvids.
Furthermore, future studies should focus on more targeted topics, such as how rapid urbanization and landscape changes affect corvid behavior and population dynamics, and how corvids adapt to climate change, particularly in extreme environments. Research could explore how shifts in climate zones impact corvid migration patterns, food availability, and habitat use. These studies would not only enhance our understanding of corvids but also contribute to broader ornithological research, aiding in the development of more effective conservation strategies. Creating bird-friendly urban spaces that support corvid populations while minimizing human-wildlife conflict is particularly important.
This study also advocates for the promotion of public bird conservation education and data collection platforms to raise awareness of the importance of protecting corvid habitats, especially in urban areas. The involvement of citizen science is crucial for enhancing data collection and fostering public engagement in bird conservation, which will support ecologists' research and expand our understanding of urban ecology. Finally, this study highlights the importance of leveraging emerging technologies in corvid and avian ecological research. In addition to existing tools like drone monitoring and GPS tracking, AI-based data analysis could provide more accurate and comprehensive insights, aiding in a deeper understanding of corvid ecology.
References
[1]. Prum, R. O., Berv, J. S., Dornburg, A., Field, D. J., Townsend, J. P., Lemmon, E. M., & Lemmon, A. R. (2015). A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature, 526(7574), 569–573. https://doi.org/10.1038/nature15697
[2]. Jetz, W., Thomas, G. H., Joy, J. B., Hartmann, K., & Mooers, A. O. (2012). The global diversity of birds in space and time. Nature, 491(7424), 444–448. https://doi.org/10.1038/nature11631
[3]. Çakar, B., Bulut, E. Ç., Kahvecioglu, O., Günay, E., Ruzhanova‐Gospodinova, I. S., & Szara, T. (2024). Bill shape variation in selected species in birds of prey. Anatomia, Histologia, Embryologia, 53(4), e13085. https://doi.org/10.1111/ahe.13085
[4]. Song, Z., Drobniak, S. M., Liu, Y., Van Schaik, C. P., & Griesser, M. (2024). Bird brains fit the bill: Morphological diversification and the evolution of avian brain size. bioRxiv, 2024-07. https://doi.org/10.1101/2024.07.02.601652
[5]. Sheard, C., Street, S. E., Evans, C., Lala, K. N., Healy, S. D., & Sugasawa, S. (2023). Beak shape and nest material use in birds. Philosophical Transactions of the Royal Society B: Biological Sciences, 378(1884), 20220147. https://doi.org/10.1098/rstb.2022.0147
[6]. Greene, E. S., Deviche, P., & Dridi, S. (2022). Reproductive behavior. In Sturkie's Avian Physiology (pp. 1111-1135). Academic Press. https://doi.org/10.1016/B978-0-12-407160-5.00030-0.
[7]. Barnagaud, J.-Y., Devictor, V., Jiguet, F., Barbet-Massin, M., Le Viol, I., & Archaux, F. (2012). Relating Habitat and Climatic Niches in Birds. PLoS ONE, 7(3), e32819. https://doi.org/10.1371/journal.pone.0032819
[8]. Guillaumet, A., & Russell, I. J. (2022). Bird Communities in a Changing World: The Role of Interspecific Competition. Diversity, 14(10), 857. https://doi.org/10.3390/d14100857
[9]. Sekercioglu, Çagan H., Wenny, Daniel G. and Whelan, Christopher J.. Why Birds Matter: Avian Ecological Function and Ecosystem Services, Chicago: University of Chicago Press, 2016. https://doi.org/10.7208/9780226382777
[10]. Chen, L., & Khanna, M. (2024). Heterogeneous and long-term effects of a changing climate on bird biodiversity. Global Environmental Change Advances, 2, 100008. https://doi.org/10.1016/j.gecadv.2024.100008
[11]. Dayer, A. A., Barnes, J. C., Dietsch, A. M., Keating, J. M., & Naves, L. C. (2020). Advancing scientific knowledge and conservation of birds through inclusion of conservation social sciences in the American Ornithological Society. The Condor, 122(4), duaa047. https://doi.org/10.1093/condor/duaa047
[12]. Stronza, A. L., Hunt, C. A., & Fitzgerald, L. A. (2019). Ecotourism for conservation?. Annual Review of Environment and Resources, 44(1), 229-253. https://doi.org/10.1146/annurev-environ-101718-033046
[13]. Döbert, T. F., Webber, B. L., Barnes, A. D., Dickinson, K. J., & Didham, R. K. (2014). Forest fragmentation and biodiversity conservation in human-dominated landscapes. In Global forest fragmentation (pp. 28-49). Wallingford UK: CABI. https://www.cabidigitallibrary.org/doi/abs/10.1079/9781780642031.0028
[14]. Drewitt, A. L., & Langston, R. H. (2008). Collision effects of wind‐power generators and other obstacles on birds. Annals of the New York Academy of Sciences, 1134(1), 233-266. https://doi.org/10.1196/annals.1439.015
[15]. Loss, S. R., Will, T., Loss, S. S., & Marra, P. P. (2014). Bird–building collisions in the United States: Estimates of annual mortality and species vulnerability. The Condor, 116(1), 8-23. https://doi.org/10.1650/CONDOR-13-090.1
[16]. Wani, H. M., Fazili, M. F., Bhat, B. A., & Bhat, R. A. Intelligence, Feeding Ecology and Scatter Hoarding Behaviour of Corvids: A review. https://envirsc.uok.edu.in/Files/ab1ac1f1-07e3-42a2-85bc-83717ef39155/Journal/ae062395-14d2-452c-b006-a3e48a30a861.pdf
[17]. McCune, K. B., Valente, J. J., Jablonski, P. G., Lee, S., & Ha, R. R. (2022). Social behavior mediates the use of social and personal information in wild jays. Scientific Reports, 12(1), 2494. https://doi.org/10.1038/s41598-022-06496-x
[18]. Palchyk, O. (2023). Overview of the Peculiarities of Social Learning of Representatives of the Genus Corvus. Path of Science, 9(1), 6010–6017. https://doi.org/10.22178/pos.89-26
[19]. Boucherie, P. H., Loretto, M.-C., Massen, J. J. M., & Bugnyar, T. (2019). What constitutes “social complexity” and “social intelligence” in birds? Lessons from ravens. Behavioral Ecology and Sociobiology, 73(1), 12. https://doi.org/10.1007/s00265-018-2607-2
[20]. Abou Zeid, F., Morelli, F., Ibáñez-Álamo, J. D., Díaz, M., Reif, J., Jokimäki, J., Suhonen, J., Kaisanlahti-Jokimäki, M.-L., Markó, G., Bussière, R., Mägi, M., Tryjanowski, P., Kominos, T., Galanaki, A., Bukas, N., Pruscini, F., Jerzak, L., Ciebiera, O., & Benedetti, Y. (2023). Spatial Overlap and Habitat Selection of Corvid Species in European Cities. Animals, 13(7), 1192. https://doi.org/10.3390/ani13071192
[21]. Jokimäki, J., Kaisanlahti-Jokimäki, M.-L., & Suhonen, J. (2022). Long-Term Winter Population Trends of Corvids in Relation to Urbanization and Climate at Northern Latitudes. Animals, 12(14), 1820. https://doi.org/10.3390/ani12141820
[22]. Benmazouz, I., Jokimäki, J., Lengyel, S., Juhász, L., Kaisanlahti-Jokimäki, M.-L., Kardos, G., Paládi, P., & Kövér, L. (2021). Corvids in Urban Environments: A Systematic Global Literature Review. Animals, 11(11), 3226. https://doi.org/10.3390/ani11113226
[23]. Szala, K., Dylewski, Ł., & Tobolka, M. (2020). Winter habitat selection of Corvids in an urban ecosystem. Urban Ecosystems, 23(3), 483–493. https://doi.org/10.1007/s11252-020-00942-2
[24]. Bragato, P. J., Spencer, E. E., Dickman, C. R., Crowther, M. S., Tulloch, A., & Newsome, T. M. (2022). Effects of habitat, season and flood on corvid scavenging dynamics in Central Australia. Austral Ecology, 47(5), 939–953. https://doi.org/10.1111/aec.13177
[25]. Cicero, C., Mason, N. A., Oong, Z., Title, P. O., Morales, M. E., Feldheim, K. A., Koo, M. S., & Bowie, R. C. K. (2022). Deep ecomorphological and genetic divergence in Steller’s Jays (Cyanocitta stelleri, Aves: Corvidae). Ecology and Evolution, 12(12), e9517. https://doi.org/10.1002/ece3.9517
[26]. Ravkin, Yu. S., Odintsev, O. A., & Bogomolova, I. N. (2023). Specificity of Studying Spatial and Typological Variations in Bird Assemblages across Certain Species Groups and Their Distribution (Using the Example of Corvidae). Contemporary Problems of Ecology, 16(1), 19–29. https://doi.org/10.1134/S1995425523010079
Cite this article
Ning,C. (2025). Corvid Intelligence and Social Complexity: An In-Depth Exploration of Adaptation Across Varied Ecological Contexts. Theoretical and Natural Science,73,64-75.
Data availability
The datasets used and/or analyzed during the current study will be available from the authors upon reasonable request.
Disclaimer/Publisher's Note
The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of EWA Publishing and/or the editor(s). EWA Publishing and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
About volume
Volume title: Proceedings of the 4th International Conference on Biological Engineering and Medical Science
© 2024 by the author(s). Licensee EWA Publishing, Oxford, UK. This article is an open access article distributed under the terms and
conditions of the Creative Commons Attribution (CC BY) license. Authors who
publish this series agree to the following terms:
1. Authors retain copyright and grant the series right of first publication with the work simultaneously licensed under a Creative Commons
Attribution License that allows others to share the work with an acknowledgment of the work's authorship and initial publication in this
series.
2. Authors are able to enter into separate, additional contractual arrangements for the non-exclusive distribution of the series's published
version of the work (e.g., post it to an institutional repository or publish it in a book), with an acknowledgment of its initial
publication in this series.
3. Authors are permitted and encouraged to post their work online (e.g., in institutional repositories or on their website) prior to and
during the submission process, as it can lead to productive exchanges, as well as earlier and greater citation of published work (See
Open access policy for details).
References
[1]. Prum, R. O., Berv, J. S., Dornburg, A., Field, D. J., Townsend, J. P., Lemmon, E. M., & Lemmon, A. R. (2015). A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature, 526(7574), 569–573. https://doi.org/10.1038/nature15697
[2]. Jetz, W., Thomas, G. H., Joy, J. B., Hartmann, K., & Mooers, A. O. (2012). The global diversity of birds in space and time. Nature, 491(7424), 444–448. https://doi.org/10.1038/nature11631
[3]. Çakar, B., Bulut, E. Ç., Kahvecioglu, O., Günay, E., Ruzhanova‐Gospodinova, I. S., & Szara, T. (2024). Bill shape variation in selected species in birds of prey. Anatomia, Histologia, Embryologia, 53(4), e13085. https://doi.org/10.1111/ahe.13085
[4]. Song, Z., Drobniak, S. M., Liu, Y., Van Schaik, C. P., & Griesser, M. (2024). Bird brains fit the bill: Morphological diversification and the evolution of avian brain size. bioRxiv, 2024-07. https://doi.org/10.1101/2024.07.02.601652
[5]. Sheard, C., Street, S. E., Evans, C., Lala, K. N., Healy, S. D., & Sugasawa, S. (2023). Beak shape and nest material use in birds. Philosophical Transactions of the Royal Society B: Biological Sciences, 378(1884), 20220147. https://doi.org/10.1098/rstb.2022.0147
[6]. Greene, E. S., Deviche, P., & Dridi, S. (2022). Reproductive behavior. In Sturkie's Avian Physiology (pp. 1111-1135). Academic Press. https://doi.org/10.1016/B978-0-12-407160-5.00030-0.
[7]. Barnagaud, J.-Y., Devictor, V., Jiguet, F., Barbet-Massin, M., Le Viol, I., & Archaux, F. (2012). Relating Habitat and Climatic Niches in Birds. PLoS ONE, 7(3), e32819. https://doi.org/10.1371/journal.pone.0032819
[8]. Guillaumet, A., & Russell, I. J. (2022). Bird Communities in a Changing World: The Role of Interspecific Competition. Diversity, 14(10), 857. https://doi.org/10.3390/d14100857
[9]. Sekercioglu, Çagan H., Wenny, Daniel G. and Whelan, Christopher J.. Why Birds Matter: Avian Ecological Function and Ecosystem Services, Chicago: University of Chicago Press, 2016. https://doi.org/10.7208/9780226382777
[10]. Chen, L., & Khanna, M. (2024). Heterogeneous and long-term effects of a changing climate on bird biodiversity. Global Environmental Change Advances, 2, 100008. https://doi.org/10.1016/j.gecadv.2024.100008
[11]. Dayer, A. A., Barnes, J. C., Dietsch, A. M., Keating, J. M., & Naves, L. C. (2020). Advancing scientific knowledge and conservation of birds through inclusion of conservation social sciences in the American Ornithological Society. The Condor, 122(4), duaa047. https://doi.org/10.1093/condor/duaa047
[12]. Stronza, A. L., Hunt, C. A., & Fitzgerald, L. A. (2019). Ecotourism for conservation?. Annual Review of Environment and Resources, 44(1), 229-253. https://doi.org/10.1146/annurev-environ-101718-033046
[13]. Döbert, T. F., Webber, B. L., Barnes, A. D., Dickinson, K. J., & Didham, R. K. (2014). Forest fragmentation and biodiversity conservation in human-dominated landscapes. In Global forest fragmentation (pp. 28-49). Wallingford UK: CABI. https://www.cabidigitallibrary.org/doi/abs/10.1079/9781780642031.0028
[14]. Drewitt, A. L., & Langston, R. H. (2008). Collision effects of wind‐power generators and other obstacles on birds. Annals of the New York Academy of Sciences, 1134(1), 233-266. https://doi.org/10.1196/annals.1439.015
[15]. Loss, S. R., Will, T., Loss, S. S., & Marra, P. P. (2014). Bird–building collisions in the United States: Estimates of annual mortality and species vulnerability. The Condor, 116(1), 8-23. https://doi.org/10.1650/CONDOR-13-090.1
[16]. Wani, H. M., Fazili, M. F., Bhat, B. A., & Bhat, R. A. Intelligence, Feeding Ecology and Scatter Hoarding Behaviour of Corvids: A review. https://envirsc.uok.edu.in/Files/ab1ac1f1-07e3-42a2-85bc-83717ef39155/Journal/ae062395-14d2-452c-b006-a3e48a30a861.pdf
[17]. McCune, K. B., Valente, J. J., Jablonski, P. G., Lee, S., & Ha, R. R. (2022). Social behavior mediates the use of social and personal information in wild jays. Scientific Reports, 12(1), 2494. https://doi.org/10.1038/s41598-022-06496-x
[18]. Palchyk, O. (2023). Overview of the Peculiarities of Social Learning of Representatives of the Genus Corvus. Path of Science, 9(1), 6010–6017. https://doi.org/10.22178/pos.89-26
[19]. Boucherie, P. H., Loretto, M.-C., Massen, J. J. M., & Bugnyar, T. (2019). What constitutes “social complexity” and “social intelligence” in birds? Lessons from ravens. Behavioral Ecology and Sociobiology, 73(1), 12. https://doi.org/10.1007/s00265-018-2607-2
[20]. Abou Zeid, F., Morelli, F., Ibáñez-Álamo, J. D., Díaz, M., Reif, J., Jokimäki, J., Suhonen, J., Kaisanlahti-Jokimäki, M.-L., Markó, G., Bussière, R., Mägi, M., Tryjanowski, P., Kominos, T., Galanaki, A., Bukas, N., Pruscini, F., Jerzak, L., Ciebiera, O., & Benedetti, Y. (2023). Spatial Overlap and Habitat Selection of Corvid Species in European Cities. Animals, 13(7), 1192. https://doi.org/10.3390/ani13071192
[21]. Jokimäki, J., Kaisanlahti-Jokimäki, M.-L., & Suhonen, J. (2022). Long-Term Winter Population Trends of Corvids in Relation to Urbanization and Climate at Northern Latitudes. Animals, 12(14), 1820. https://doi.org/10.3390/ani12141820
[22]. Benmazouz, I., Jokimäki, J., Lengyel, S., Juhász, L., Kaisanlahti-Jokimäki, M.-L., Kardos, G., Paládi, P., & Kövér, L. (2021). Corvids in Urban Environments: A Systematic Global Literature Review. Animals, 11(11), 3226. https://doi.org/10.3390/ani11113226
[23]. Szala, K., Dylewski, Ł., & Tobolka, M. (2020). Winter habitat selection of Corvids in an urban ecosystem. Urban Ecosystems, 23(3), 483–493. https://doi.org/10.1007/s11252-020-00942-2
[24]. Bragato, P. J., Spencer, E. E., Dickman, C. R., Crowther, M. S., Tulloch, A., & Newsome, T. M. (2022). Effects of habitat, season and flood on corvid scavenging dynamics in Central Australia. Austral Ecology, 47(5), 939–953. https://doi.org/10.1111/aec.13177
[25]. Cicero, C., Mason, N. A., Oong, Z., Title, P. O., Morales, M. E., Feldheim, K. A., Koo, M. S., & Bowie, R. C. K. (2022). Deep ecomorphological and genetic divergence in Steller’s Jays (Cyanocitta stelleri, Aves: Corvidae). Ecology and Evolution, 12(12), e9517. https://doi.org/10.1002/ece3.9517
[26]. Ravkin, Yu. S., Odintsev, O. A., & Bogomolova, I. N. (2023). Specificity of Studying Spatial and Typological Variations in Bird Assemblages across Certain Species Groups and Their Distribution (Using the Example of Corvidae). Contemporary Problems of Ecology, 16(1), 19–29. https://doi.org/10.1134/S1995425523010079