







1. Introduction
Throughout history, changes in climate in their living environments have led some animals to evolve a mysterious and fascinating structure, such as a false thumb. The possession of a false thumb is almost the most conspicuous cases of convergence in vertebrates [1]. It has been used to adapt to changes caused by dietary changes or the construction of living burrows, such as grasping or enlarging the surface of the hand to allow for patting and pressing soil or to make locomotion easier, and to reduce the need for specialisation of the hand due to the acquisition of food in order to re-satisfy the need for walking speed [1,2]. But it is not really a thumb in the traditional cognitive and anatomical sense. The false thumb of pandas, both giant pandas and red pandas, is a radial sesamoid bone; the sesamoid bone is one of the carpal bones, but usually serves important functions opposite the thumb, such as grasping and fine handling of food, among other actions [1-4], a section that will be referred to in the following section.
In a study conducted by Morales et al. 2005 that false thumbs found in Simocyon batalleri (extinct), a close relative of the Miocene red panda, suggests that trait was inherited from a common ancestor in the red panda family (Ailuridae) [5]. The false thumb likely originated as an aid to arboreal locomotion. In the context of panda evolution and the development of false thumbs, it showcases how nature can creatively utilize pre-existing anatomical features to meet novel demands. This reflects the nature of evolutionary opportunism. The false thumb was used as an aid in adapting to arboreal locomotion in panda-originating species such as Simocyon batalleri and was later used to manipulate bamboo in red pandas and giant pandas [5]. By repurposing this skeleton for grasping and handling food.
Evolutionary randomness as well as evolutionary opportunism is a good entry point for conjecture as to why these animals chose to evolve the sesamoid as a thumb-assisted grasping and stable genetic retention to this day, which will be mentioned later on in this paper in the suggestion section.
In most of the existing articles on false thumb, each animal species is discussed separately. Few existing papers have analysed animals that have evolved false thumbs together and found their intrinsic connection from an evolutionary biology perspective. This study outlines the function (evolutionary intent) of false thumbs in several animals that have been found to possess false thumbs. Furthermore, this study describes about evolutionary conjectures linking the well-researched giant panda to animals with false thumb but to be studied. In attempt to bring new ideas up for research, this literature discusses the evolutionary and developmental history of the giant panda in whilst accordance to Darwinian evolutionary theory.
2. Analysis of the function and morphology of the false thumb in each animal
2.1. Panda’s false thumb
The giant panda has six fingers. But anatomically, the sixth finger is not really a thumb. Giant panda’s false thumb evolved from the radial sesamoid bone [1,5]. Giant pandas have adapted to the environment by making the radial sesamoid bone larger and more medial, with the cartilage curved apically. With the articular surface of the sesamoid bone covered with cartilage, the apical curvature allows the mabductor digiti Ibrevie to have a more complete bony attachment. Connecting first with the metacarpal, then lead to higher efficiency in the pincer-like mechanism the pulse thumb [4,5]. This evolutionary change has given the giant panda a more efficient grip, evolving the radial sesamoid into a grip structure that can handle food (bamboo leaves and stems) more efficiently [6].
The red panda also has a false thumb. Despite their evolutionary division into two species (Giant pandas belong to the order Carnivora, family Ursidae, subfamily Giant Panda, genus Giant Panda. The scientific name of this species is Ailuropoda melanoleuca, which translates to "black and white catsfoot". Although red pandas, similar to giant pandas, are classified within the order Carnivora, they belong to distinct families, subfamilies, and genera. It belongs to the family Ailuridae, subfamily Ailuridae and genus Ailurus. Its scientific name is Ailurus fulgens, which means "shiny catsfoot"), both achieve their grasping function by enlarging the radial raphe bone highlighting their remarkable similarity in grasping mechanisms.
However, there are still some segmented functional differences between them. The red panda is a small arboreal animal, its false thumb is used not only for feeding unlike the giant panda which uses the false thumb mainly for delicate handling of bamboo leaves and stems, but also for climbing [6]. Red panda’s false thumb has evolved more to adapt to grasping slender branches rather than extensive manipulation tasks. The two pandas differ significantly in their muscle insertion sites. Muscle insertions in red pandas include the adductor muscle on the first metacarpal bone and the palmaris longus muscle on the radial raphe, which enhance their upward and inward movements when climbing and moving in arboreal environments. Giant pandas, on the other hand, are terrestrial animals, which causes them to have different muscle insertion sites. This makes the evolution of the false thumb of the red panda a little less developed compared to the giant panda [5]. Compared to the giant panda, the false thumb of the red panda is relatively small and less hypertrophied. The false thumb has fewer direct connections to the carpal bones and a different structural orientation. This results in different functional aspects of the radial sesamoid process.
It can find that this glaring disparity in the two graphs below from Edwin Dickinson's study and Gould's book, respectively shown in Figure 1 and Figure 2.
|
| Figure 1. Red panda [5] | Figure 2. Giant panda [3] |
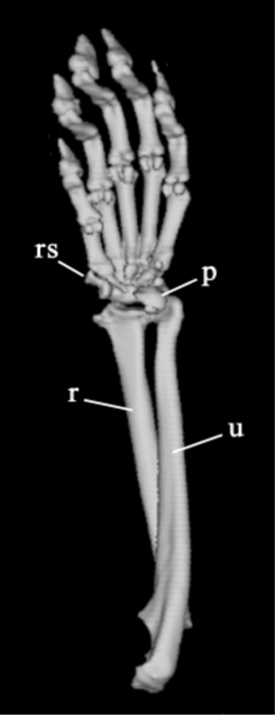
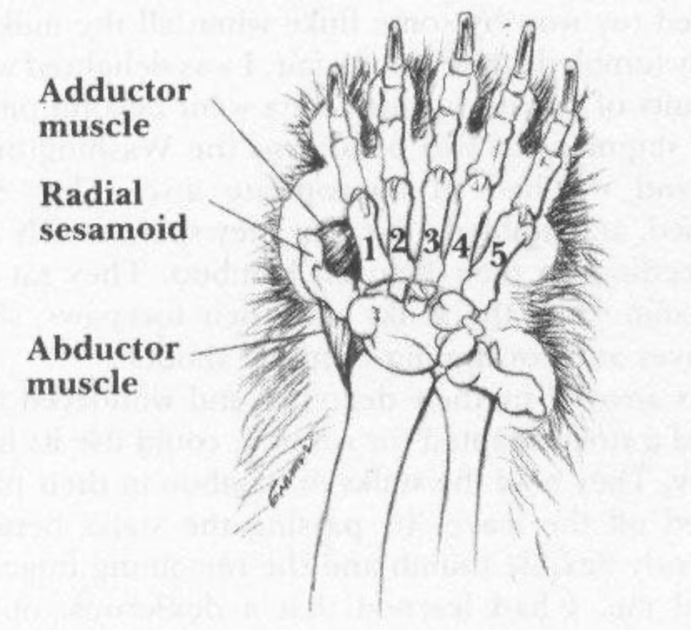
2.2. Mole’s false thumb
Mole also has a false thumb. Same to the giant panda and red panda, the mole’s false thumb is also evolved by the sesamoid bone. Their palms are significantly widened by the false thumb. So that moles can use their widened palms to pat down loose soil against the tunnel walls instead of bothering to carry soil to the surface [7, 8]. (This evolved to allow moles to consume less energy and reduce the pressure on moles to survive). As mentioned in Yi-fen's et al. article, the false thumb is capable of supporting a significant portion of the mole rat's body weight, akin to the role of the radial sesamoid bone in elephants, especially during changes in foot posture. This weight-bearing function allows mole rats to maintain stability and efficient locomotion while digging burrows and living in burrows with complex structures [9]. The experiment in Yi-Fen's article also mentions that during walking, only the "false thumb" and "Digit I" are in contact with the ground." The "false thumb" touches the ground first, and together with "Digit I"(it’s a structure homologous with human’s thumb), which then touches the ground, they form a stable triangular support structure. This contact helps the mole to remain stable and balanced during movement. We can see this situation in Figure 3 from Yi-Fen’s et al. study [7].
Figure 3. Mole walking [7]
2.3. Aye-aye’s false thumb (Daubentonia madagascariensis)
Aye-aye's false thumb also evolved from the radial sesamoid bone, like the panda's, by enlarging the sesamoid bone to adapt to new survival needs brought about by environmental changes. The false thumb has muscular attachments from three muscles, enabling abduction, adduction, and opposition. However, it is functionally very different from the panda.
Aye-aye evolved the false thumb not for gripping, but to compensate for the overspecialisation of the fingers in non-gripping functions [10]. Aye-aye has highly specialized third digit and fourth digit, and Tomáš Jůnek et al. found that the third finger was usually used for percussion, cleaning teeth, and detecting nectar. The fourth digit is more commonly used to eat pineapple nectar than the third finger. In the detection of invertebrates in plant cortex and the trunk of dead trees, the two fingers were used in proportion to each other. Whereas the two fingers are assigned to distinct tasks while extracting fruit from coconuts, the third digit demonstrates exceptional skill in executing tasks that require high degrees of agility, dexterity, and precision, in contrast to the fourth digit, which appears to be more proficient in managing tasks that necessitate strength, a scooping action, and deep contact [11]. Specifically, aiding in the aye-aye's unique "tap foraging" practices. The aye-aye's false thumb allows the aye-aye to get a better grip on relatively small items, making the aye-aye more efficient and stable when using them. False thumbs allow aye-aye to improve palm dexterity and grip, enabling them to perform unique foraging methods such as knocking on wood to locate their prey wood-boring larvae, extracting food, and exploring their environment with greater precision. The false thumb allows aye-aye to use tools with dexterity and to locate in the forest [10].
2.4. Spectacled bear’s false thumb (Tremarctos ornatus)
The spectacled bear’s false thumb also evolved from radial sesamoid. The false thumb allows spectacled bears to access and process food more efficiently. Spectacled bears can use their false thumbs to process, extract and ingest specific parts of plants (e.g. bamboo, pineapple [12]) or other food sources that have nutritional value or are easier to digest. This improves their feeding, obtains energy efficiency, and ensures that they are able to meet their survival needs. In addition, spectacled bears are arboreal and false thumbs help them climb in trees. The enlarged radial sesamoid bone provides additional support and grip, enhancing the spectacled bear's ability to move in arboreal environments. This adaptation is particularly advantageous for accessing tree food sources or avoiding predators [13].
The bear false thumb serves a similar purpose as those in the pandas, moreover the false thumb in the bear looks like a combination of the red and giant pandas’ false thumb. Spectacled bears, like giant pandas, are a species that typically possess false thumbs [14].
3. Factors affecting the development of false thumbs
3.1. Genetic factors
The formation of the false thumb gene may be the result of mutations in one or more of the genes responsible for the new phenotype. Since these mutations give animals with false thumbs better access to processed food for survival, these mutations provide an evolutionary advantage and, according to Darwin's doctrine of natural selection, these animals are therefore better able to find mates and become stronger as a result of the mutations, leaving the original unaltered genes progressively at a disadvantage. Conversely, genes that are advantageous, i.e.genes that form the false thumb phenotype, can be passed on to the next generation, gradually creating a stable phenotype and eliminating those of the same kind that do not have the mutated genes.
3.2. Environmental factors
The earliest known fossil of a giant panda with false thumb features was found at the late Miocene site of Shuitangba in Yunnan, China [6]. During the late Miocene epoch, characterized by fluctuations in global climate, significant fossil discoveries were made, including changes in temperature and precipitation patterns. These fluctuating climates led to more pronounced seasons, promoting environmental diversity and heterogeneity in terrestrial ecosystems. More new plants and animals appeared. Different regions presented a variety of habitats, including forests, grasslands, wetlands and savannas [15]. This means that the diet of primitive pandas surviving in forests would have changed as a result, and having a false thumb would have been more favourable for survival.
4. Suggestion
There have study already found that the "thumb" has not increased in size since the late Miocene, because when walking in plantigrade posture, the "thumb" had to be used to carry its own weight as well as to maintain balance, like an elephant, and could not be specialised like mole's second and third digits [6]. An interesting question that came up while reading the paper was why all these animals with false thumbs chose to evolve the radial seed bone instead of modifying their existing fingers.
Existing fingers already have specialised uses, such as Mole's highly specialised second and third fingers, and there is no selective pressure to allow false thumbs to evolve from existing fingers [7]. And if the choice had been to mutate the existing fingers, it could instead have made the aforementioned false thumbs dramatically less capable of hunting and surviving. Because they already had a well-established system and balance of hunting through existing fingers, mutating existing fingers would not be advantageous in natural selection.
From an evolutionary opportunistic point of view, the opportunity cost of mutating a sesamoid already in the position of a thumb is lower compared to the opportunity cost of mutating an already mature finger, even though the thumb evolved in this way would look a little rough. False-thumbed animals would have preferred the convenience of short-term predation, even though it may be that in the future more refined ways of handling food could give them a greater survival advantage. Pandas, for example, who are carefully cared for in zoos, no longer seem to have to worry that over-specialising their thumbs will slow them down, prevent them from catching up with their prey or escaping their natural predators and losing their lives.
Genetically speaking, there may be genetic limitations that have not yet been discovered, which explains why the genome has not changed to give the existing fingers a new phenotype. There is no change in the genetic code that would allow for the modification or evolution of existing fingers. The current functioning of giant panda fingers allows them to adequately perform their daily tasks, so there is no anatomically imminent selection pressure. This could explain why giant pandas did not evolve their existing fingers, but only a small sesamoid bone, a false thumb, to help them get a slightly better grip. They don't need to specialise in more flexible thumbs.
The absence of the false thumb from fully evolving into a complete finger can primarily be attributed to the presence of limiting factors, with weight-bearing being one of the primary constraints. The development of the false thumb has reached a plateau, resulting in a small, flat structure that remains barely distinguishable from the palmar surface.
Although all studies point to Darwinian evolution as explaining the advantageous aspects of false thumbs, there are still a number of factors that cannot be explained, including the inability to fully explain the genetic mechanisms behind false fingers and the role of environmental factors that influence the growth of these structures. And it's difficult avoid Gettier's paradox when it comes to the analysis of the section on factors affecting false thumbs. Furthermore, the evolutionary significance of these false thumb is unclear, and it is not known whether they actually improve the probability of survival or reproductive success.
5. Conclusion
In summary, this study analyses the function and anatomy of false thumbs in animals that have been identified so far and possess false thumb structures. The environmental context of the earliest known fossil panda false thumb structure is analysed. This study analysed the functional, structural and morphological similarities and differences between the false thumbs of giant pandas, red pandas, mole rats, ye-aye and spectacled bears in an attempt to find out the intrinsic similarities and logic behind the phenomenon of "different paths leading to the same place". Based on Darwinian evolutionary theory, as well as philosophically orientated conjectures, a plausible explanation for the origin of the false thumb structure is provided. Speculations are made as to why these false thumbs chose to evolve the radial sesamoid as a false thumb rather than mutate the original finger structure. This study converges to summaries current research on the direction of animals with false thumbs adopts some philosophical perspectives to explain the formation of false thumbs, and hopefully provides a new way of thinking for researchers in the same field.
References
[1]. Salesa, M. J. et al. (2006). Anatomy of the “false thumb” of Tremarctos ornatus (Carnivora, Ursidae, Tremarctinae): phylogenetic and functional implications.
[2]. Salesa, M. J. at al. (2006). Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas. Proceedings of the National Academy of Sciences, 103(2), 379-382.
[3]. Gould, S.J. Te Panda’s Tumb,( 1982) More Refections in Natural History paperback edn. (W. W. Norton & Company, New York.
[4]. Abella, J. et al. (2015). Tracing the origin of the panda’s thumb. The science of Nature, 102, 1-13.
[5]. Antón, M. et al. (2006). Implications of the functional anatomy of the hand and forearm of Ailurus fulgens (Carnivora, Ailuridae) for the evolution of the ‘false‐thumb’in pandas. Journal of anatomy, 209(6), 757-764.
[6]. Wang, X. et al. (2022). Earliest giant panda false thumb suggests conflicting demands for locomotion and feeding. Scientific reports, 12(1), 10538.
[7]. Lin, Y. F., Konow, N., & Dumont, E. R. (2019). How moles walk; it's all thumbs. Biology Letters, 15(10), 20190503.
[8]. Lin, Y. F., Konow, N., & Dumont, E. R. (2019). How moles destroy your lawn: the forelimb kinematics of eastern moles in loose and compact substrates. Journal of Experimental Biology, 222(4), jeb182436.
[9]. Yalden, D. W. (1966). The anatomy of mole locomotion. Journal of Zoology, 149(1), 55-64.
[10]. Lhota, S., Jůnek, T. et al. (2008). Specialized use of two fingers in free‐ranging aye‐ayes (Daubentonia madagascariensis). American Journal of Primatology: Official Journal of the American Society of Primatologists, 70(8), 786-795.
[11]. Hartstone‐Rose, A. et al. (2020). A primate with a Panda's thumb: The anatomy of the pseudothumb of Daubentonia madagascariensis. American journal of physical anthropology, 171(1), 8-16.
[12]. Peyton, B. (1980). Ecology, distribution, and food habits of spectacled bears, Tremarctos ornatus, in Peru. Journal of Mammalogy, 61(4), 639-652.
[13]. Salesa, M. J. et al. (2006). Anatomy of the “false thumb” of Tremarctos ornatus (Carnivora, Ursidae, Tremarctinae): phylogenetic and functional implications.
[14]. Vela-Vargas, I. M. et al. (2021). Tremarctos ornatus (Carnivora: Ursidae). Mammalian Species, 53(1006), 78-94.
[15]. Jablonski, N. G. et al. (2014). The site of Shuitangba (Yunnan, China) preserves a unique, terminal Miocene fauna. Journal of Vertebrate Paleontology, 34(5), 1251–1257Reference lists.Caplar R and Kulisic P 1973 Proc. Int. Conf. on Nuclear Physics (Munich) vol 1 (Amsterdam:North-Holland/American Elsevier) p 517
Cite this article
Qiu,S. (2024). Analyses based on the function of false thumb in animals possessing false thumbs. Theoretical and Natural Science,58,41-46.
Data availability
The datasets used and/or analyzed during the current study will be available from the authors upon reasonable request.
Disclaimer/Publisher's Note
The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of EWA Publishing and/or the editor(s). EWA Publishing and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
About volume
Volume title: Proceedings of the 4th International Conference on Biological Engineering and Medical Science
© 2024 by the author(s). Licensee EWA Publishing, Oxford, UK. This article is an open access article distributed under the terms and
conditions of the Creative Commons Attribution (CC BY) license. Authors who
publish this series agree to the following terms:
1. Authors retain copyright and grant the series right of first publication with the work simultaneously licensed under a Creative Commons
Attribution License that allows others to share the work with an acknowledgment of the work's authorship and initial publication in this
series.
2. Authors are able to enter into separate, additional contractual arrangements for the non-exclusive distribution of the series's published
version of the work (e.g., post it to an institutional repository or publish it in a book), with an acknowledgment of its initial
publication in this series.
3. Authors are permitted and encouraged to post their work online (e.g., in institutional repositories or on their website) prior to and
during the submission process, as it can lead to productive exchanges, as well as earlier and greater citation of published work (See
Open access policy for details).
References
[1]. Salesa, M. J. et al. (2006). Anatomy of the “false thumb” of Tremarctos ornatus (Carnivora, Ursidae, Tremarctinae): phylogenetic and functional implications.
[2]. Salesa, M. J. at al. (2006). Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas. Proceedings of the National Academy of Sciences, 103(2), 379-382.
[3]. Gould, S.J. Te Panda’s Tumb,( 1982) More Refections in Natural History paperback edn. (W. W. Norton & Company, New York.
[4]. Abella, J. et al. (2015). Tracing the origin of the panda’s thumb. The science of Nature, 102, 1-13.
[5]. Antón, M. et al. (2006). Implications of the functional anatomy of the hand and forearm of Ailurus fulgens (Carnivora, Ailuridae) for the evolution of the ‘false‐thumb’in pandas. Journal of anatomy, 209(6), 757-764.
[6]. Wang, X. et al. (2022). Earliest giant panda false thumb suggests conflicting demands for locomotion and feeding. Scientific reports, 12(1), 10538.
[7]. Lin, Y. F., Konow, N., & Dumont, E. R. (2019). How moles walk; it's all thumbs. Biology Letters, 15(10), 20190503.
[8]. Lin, Y. F., Konow, N., & Dumont, E. R. (2019). How moles destroy your lawn: the forelimb kinematics of eastern moles in loose and compact substrates. Journal of Experimental Biology, 222(4), jeb182436.
[9]. Yalden, D. W. (1966). The anatomy of mole locomotion. Journal of Zoology, 149(1), 55-64.
[10]. Lhota, S., Jůnek, T. et al. (2008). Specialized use of two fingers in free‐ranging aye‐ayes (Daubentonia madagascariensis). American Journal of Primatology: Official Journal of the American Society of Primatologists, 70(8), 786-795.
[11]. Hartstone‐Rose, A. et al. (2020). A primate with a Panda's thumb: The anatomy of the pseudothumb of Daubentonia madagascariensis. American journal of physical anthropology, 171(1), 8-16.
[12]. Peyton, B. (1980). Ecology, distribution, and food habits of spectacled bears, Tremarctos ornatus, in Peru. Journal of Mammalogy, 61(4), 639-652.
[13]. Salesa, M. J. et al. (2006). Anatomy of the “false thumb” of Tremarctos ornatus (Carnivora, Ursidae, Tremarctinae): phylogenetic and functional implications.
[14]. Vela-Vargas, I. M. et al. (2021). Tremarctos ornatus (Carnivora: Ursidae). Mammalian Species, 53(1006), 78-94.
[15]. Jablonski, N. G. et al. (2014). The site of Shuitangba (Yunnan, China) preserves a unique, terminal Miocene fauna. Journal of Vertebrate Paleontology, 34(5), 1251–1257Reference lists.Caplar R and Kulisic P 1973 Proc. Int. Conf. on Nuclear Physics (Munich) vol 1 (Amsterdam:North-Holland/American Elsevier) p 517